

اگر به یک وب سایت یا فروشگاه رایگان با فضای نامحدود و امکانات فراوان نیاز دارید بی درنگ دکمه زیر را کلیک نمایید.
ایجاد وب سایت یا ازمایشگاه تکنیک پالس شبیهسازی با proteus
ازمایشگاه تکنیک پالس شبیهسازی با proteus تجزیه و تحلیل نقاشی مدرسه آتن
تجزیه و تحلیل نقاشی مدرسه آتن مقاله مدیریت سازمان های ورزشی
مقاله مدیریت سازمان های ورزشی كاربرد استخراج با سيال فوق بحراني در صنايع غذايي
كاربرد استخراج با سيال فوق بحراني در صنايع غذايي روش های نوین تجارت و تجارت الکترونیکی فرش دستباف
روش های نوین تجارت و تجارت الکترونیکی فرش دستباف تحقیق درباره ی Access
تحقیق درباره ی Access تحقیق كارشناسي ارشد عوامل موثر در روسپي گري فحشا
تحقیق كارشناسي ارشد عوامل موثر در روسپي گري فحشا هنر و گرافیک
هنر و گرافیک بررسی و طراحی نرم افزار حسابداری و مالی
بررسی و طراحی نرم افزار حسابداری و مالی تحقیق درباره ی هیدروژناسیون
تحقیق درباره ی هیدروژناسیون نقش و کاربرد انرژی هسته ای در کشاورزی
نقش و کاربرد انرژی هسته ای در کشاورزی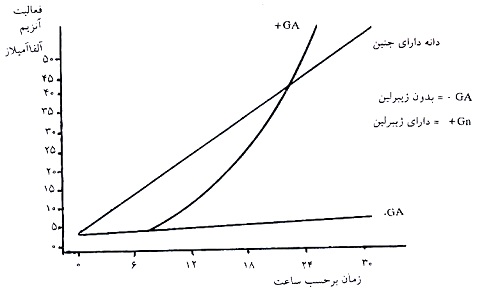 تحقیق تنظيم كننده هاي رشد گياهي (هورمونهاي گياهي)
تحقیق تنظيم كننده هاي رشد گياهي (هورمونهاي گياهي) فروش فیلتر بورسی استریکلی فقط 75 هزار تومان
فروش فیلتر بورسی استریکلی فقط 75 هزار تومان کسب درآمد اینترنتی 300000 تومان در خانه در کمتر از 30 دقیقه
کسب درآمد اینترنتی 300000 تومان در خانه در کمتر از 30 دقیقه کسب درآمد روزانه حداقل یک میلیون تومان ! کاملا حلال و واقعـی !!
کسب درآمد روزانه حداقل یک میلیون تومان ! کاملا حلال و واقعـی !! مجموعه ی آموزش تعمیر لامپ کم مصرف (از مبتدی تا پیشرفته)
مجموعه ی آموزش تعمیر لامپ کم مصرف (از مبتدی تا پیشرفته) کسب و کار اینترنتی در منزل
کسب و کار اینترنتی در منزل آموزش برنامه نویسی آردوینو
آموزش برنامه نویسی آردوینو آموزش رایگان کسب درآمد از سایت الیمپ ترید ( olymp trade )
آموزش رایگان کسب درآمد از سایت الیمپ ترید ( olymp trade ) دانلود نمونه فاکتور آماده با فرمت ورد - اکسل و عکس
دانلود نمونه فاکتور آماده با فرمت ورد - اکسل و عکس آموزش ساخت بازی بدون دانش برنامه نویسی و طراحی سه بعدی مبتدی تا پیشرفته با نرم افزار
آموزش ساخت بازی بدون دانش برنامه نویسی و طراحی سه بعدی مبتدی تا پیشرفته با نرم افزار اموزش کسب درامد از اینترنت روزانه ۳میلیون تومان تضمینی و تست شده
اموزش کسب درامد از اینترنت روزانه ۳میلیون تومان تضمینی و تست شده درامدزایی در خواب! (تعجب نکنید! بخوانید)
درامدزایی در خواب! (تعجب نکنید! بخوانید) مدار داخلی واکی تاکی(اموزش ساخت)
مدار داخلی واکی تاکی(اموزش ساخت) کتاب افزایش ممبر کانال تلگرام
کتاب افزایش ممبر کانال تلگرام دانلود100% رایگان نرم افزار تبلیغات در تلگرام + آموزش کامل و فیلم آموزشی
دانلود100% رایگان نرم افزار تبلیغات در تلگرام + آموزش کامل و فیلم آموزشی روش اصلی موفقیت در کنکور و آزمون ها(پزشکی، حقوق، مهندسی، نمونه و تیزهوشان) با پکیج کنکورپلاس
روش اصلی موفقیت در کنکور و آزمون ها(پزشکی، حقوق، مهندسی، نمونه و تیزهوشان) با پکیج کنکورپلاسهورمونهاي گياهي
تنظيم كننده هاي رشد گياهي........................................................................................................... 2
بيان عمومي.................................................................................................................................... 3
انتقال اكسين در گياه....................................................................................................................... 6
نقش اكسين در بكرزايي............................................................................................................... 11
ژيبرلين ها.................................................................................................................................... 12
انتقال ژيبرلين در گياه :............................................................................................................... 15
اثرات فيزيولژيكي ژيبرلين.......................................................................................................... 15
سيتوكينين ها :.............................................................................................................................. 16
بازدارنده هاي رشد گياهي........................................................................................................... 20
آبسزيك اسيد................................................................................................................................. 21
انتقال آبسيزيك اسيد...................................................................................................................... 23
اتيلن.............................................................................................................................................. 25
اثرات فيزيولژيكي اتيلن در گياه................................................................................................... 26
دوره زماني توليد اتيلن و فعاليت ACC سنتاز بعد از زخم ميوه سبز گوجه فرنگي.................... 30
درك اين مطلب كه گياهان چگونه رشد مي كنند نه تنها براي دست اندركاران علوم گياهي بلكه براي عموم بيولژيست ها قابل اهميت و توجه است. واژه هورمن (Hormone) ريشه يوناني دارد اين كلمه به معني محرك است كه نخستين بار در مبحث فيزيولوژي جانوري به كار برده شد. هنگامي كه مشخص شد در گياهان نيز مواد محرك رشد وجود دارند واژه هورمون تحت عنوان هورمونهاي گياهي (Phytohormone) به كار گرفته شد بررسيهاي بعدي نشان داد كه گروهي از مواد در گياهان داراي نقش تنظيم كنندگي هستند به طوري كه برخي از آنها اثرات تحريك كنندگي و بعضي اثرات بارندگي دارند اين مواد مجموعا تحت عنوان تنظيم كننده هاي گياهي يا plant growth regulators (pgr) ناميده مي شوند .
در تقسيم بندي اوليه مواد تنظيم كننده رشد درون زاي گياهان به پنج گروه تقسيم شدند كه شامل اكسين ها auxins ژيبرلين ها (ga) Gibberellins ، سيتوكينينها (CK) cytokininsآبسيزيك اسيد (aba) abscisic acid و اتيلن Etylene مي باشند .
اكسين ها و ژيبرلينها به عنوان تنظيم كننده هاي طويل شدن سلولي،سيتوكينين ها به عنوان تنظيم كننده تقسيم سلولي، آبسيزيك اسيد به عنوان يك ماده با اثرات بازدارندگي واتيلن به عنوان يك ماده فرارموثردر فرآيندهاي رشد ونمو شناخته
شده اند.مباحث مربوط به تنظيم كننده هاي رشد گياهي نخستين بار در نيمه دوم قرن نوزدهم به طور جدي در فيزيولژي گياهي وارد شد .
در سال 1880 داروين در مورد اثرات نور بر گياهان مطالعاتي انجام داد و مشاهده كرد هنگامي كه نور از يك جهت بر قسمت راسي ساقه مي تابد گياه نسبت به جهت تابش نور خميدگي پيدا مي كند درحالي كه اگر قسمتي از راس ساقه گياه كه در معرض نورقرارگرفته است به وسيله كاغذ پوشانده شود خميدگي ايجاد نمي گردد اين پژوهشهاسرآغاز اوليه در ارتباط با تنظيم كننده هاي رشد گياهي بود تنظيم كننده هاي رشد گياهي مواد آلي هستند كه متنوع بوده و از نظر مكانيسم عمل با ويتامين ها ، آنزيم ها و كوآنزيم ها تفاوت دارند امروزه برخي از مواد تنظيم كننده رشد گياهي به طور صنعتي توليد مي شوند ماده اي از نظر فيزيولوژيكي تنظيم كننده رشد طبيعي در گياهان محسوب مي شود كه حداقل داراي سه ويژگي زير باشد :
بنابراين تنظيم كننده رشد گياهي ماده اي است كه در بخشي از پيكر گياه
ساخته شده و در همان محل يا پس از انتقال در محل درگيري از بدن گياه اثرات محسوسي را ايجاد مي كند. در بدن جانوران برخي هورمونها در غددي كه ترشحات خود را مستقيما به درون مي ريزند (غدد درون ريز ) ساخته مي شوند كه از آن جمله مي توان تيروكسين از غده تيروئيد و انسولين از پانكراس و .... را نام برد در گياهان تركيبات مترادفي كه از نظر شيميايي متفاوت هستند توليد شوند.اختصاصي بودن اثر تنظيم كننده هاي رشد گياهي مانند آنچه در جانوران وجود دارد در گياهان ديده
نمي شود . اين تفاوتها احتمالا به علت محدود بودن تمايز در گياهان است .
اكسين ها
در سال 1880 داروين بررسيهايي را در اتباط با اثر تابش نور بر خيمدگي در بخشهاي راسي گياهي انجام داد . وي در اين بررسيها از كولئوپتيل و فالاريس استفاده كرد . پس از داروين فيتينگ در سال 1907 با بررسيهايي كه با كولئوپتيل جو دو سر انجام داد مشاهده كرد كه اگر قسمت راسي كولئوپتيل را قطع كرده و دوباره آن را سر جاي خود بگذارد و سپس آن را در معرض تابش نور يك جانبه قرار دهد راس كولئوپتيل در ادامه مراحل رشد خود به طرف جهت تابش نور خميدگي پيدا مي كند به اين ترتيب مشخص شد كه انتقال محرك به وسيله بريدگي ايجاد شده در كولئوپتيل قطع نشده است . اين بررسي در سالهاي 1913-1910 بوسيله بوريسن جانسن دنبال شد و نشان داد كه اگر پس از قطع راس كولئوپتيل بين دو قسمت جدا شده ( راس و پايه ) يك لايه ژلوزياميكا قرار داده شود و تحت تاثير نور يك جانبه واقع گردد باز هم خميدگي به طرف جهت تابش نور ايجاد مي شود در سال 1919 پل راس كولئوپتيل را قطع كرده و آن را طوري روي پايه قرار داد كه راس و پايه در يك امتداد نبودند و اين بار نيز مشاهده شد كه تحت تاثير تابش يك سويه نور خميدگي ايجاد مي شود . پل با تكرار بررسيهاي جانسن –بويسن نشان داد كه در اين موارد يك ماده محرك يا هورمون دخالت دارد در تجربيات ديگري پل بين دو قسمت قطع شده راس كولئوپتيل صفحه پلاتيني قرار داد و مشاهده كرد كه محرك از آن عبور نمي كند و لذا دريافت كه پديده الكتريكي در اين مورد دخالت ندارد و سرانجام در آزمايش ديگري پل بين دو بخش قطع شده راس كولئوپتيل لايه اي از كره كاكائو قرار داد كه اين بار نيز محرك عبور نمي كند و معلوم شد كه ماده محرك در چربي محلول نبوده و محلول در آب است زيرا از كره كاكائو عبور نكرده ولي از ژلاتين و ميكا عبور كرده است .
در سالهاي 1925-1923 soding راس كولئوپتيل را قطع كرده و دوباره آن را بر جاي خود قرار داد و مشاهده كرد كه رشد خطي انجام مي شود و اگر راس كولئوپتيل جدا گردد و در محل خود قرار داده نشود رشد آن صورت نمي گيرد .
در سال 1928 ونت حدس زد كه در راس كولئوپتيل ماده اي است كه محرك رشد بوده و در خميدگي آن موثر مي باشد نامبرده تعدادي مكعب ژلوژ تهيه كرده و راس كولئوپتيل يولاف جدا شده را بر روي دايره هاي ژلوزي قرار داد و آنها را در معرض نور گذاشت وي اظهار داشت اگر در راس كولئوپتيل ماده اي باشد بايد به ژلوز منتقل گرد . ونت سپس مكعبهاي ژلوز را از راس كولئوپتيل قطع شده جدا كرد و آنها را روي پايه اي از كولئوپتيل بدون راس قرار داد و مشاهده كرد كه رشد كولئوپتيل بدون راس صورت مي گيرد و در اثر يك جهت بودن ژلوز بر روي كولئوپتيل بدون راس كولئوپتيل راس خميدگي نيز ايجاد مي شود به اين ترتيب ونت آزمايش مشهور خود را انجام داد و توانست ماده محرك را به ژلوز وارد كند و موفق به استخراج ماده محرك گردد. اين آزمايش به شيميدانهاي هلندي كوگل و هاگن-اسميت اين امكان را داد كه در سال 1931 روش جداسازي و تخليص اين ماده را بدست آورند .
در سال 1934 كوگل ماده موجود در راس كولئوپتيل را مشخص كرد و آنرا اكسين ناميد. اكسين يك واژه يوناني است و از Auxe به معني بزرگ شدن يا رشد گرفته شده است به اين ترتيب ماده محرك رشد در گياهان به نام اكسين شناخته شد و از اين به بعد تجربيات بسيار زيادي در ارتباط با استخراج و شناسايي اين گروه از مواد در گياهان مختلف حتي ادرار پستانداران صورت گرفته است اكسين ها گروهي از هورمونهاي گياهي هستند كه باعث طويل شدن سلولهاي گياهي مي گردند اين مواد طيف گسترده اي را ازنظر واكنشهاي رشد و نموي در گياهان سبب مي شوند .واژه اكسين يك اصطلاح ژنريك است وبه تعدادي از مواد طبيعي اطلاق مي گردد.مهمترين و فراوانترين اكسين ها در گياهان اندول استيك اسيد است كه بارزترين اثرات را در اين گروه از هورمونهاي گياهان نشان مي دهد. مهمترين اثراتي كه تاكنون به هورمونهاي اكسين نسبت داده شده است عبارتند از بزرگ شدن سلول گياهي ، طويل شدن ساقه گياه ، توليد ريشه ، توليدآوند چوبي ، افزايش رشد جوانه راسي ، جلوگيري از رشد جوانه جانبي ، تشكيل ميوه ، بزرگ شدن ميوه ، تشكيل گرهك در ريشه گياهاني كه داراي باكترهاي تثبيت كننده نيتروژن هستند ، جلوگيري از ريزش برگ، بيو سنتز پروتئين ، بيو سنتز RNA و اثرات ديگر .
مهمترين اكسين ها اندول استيك اسيد است كه ساختمان شيميايي آن به صورت زير است
اين ماده در گياهان فراوان بوده و در برخي از قارچها نيز وجود دارد
هورمون اكسين پس از تشكيل گياه ممكن است اثرات خود را در همان محل توليد اعمال نمايد و يا در گياه منتقل شده و در ناحيه اي غير از محل تشكيل اثرات خود را آشكار سازد . اندول استيك اسيد كه توليد مي شود نه تنها كمتر به صورت آزاد درگياه باقي مي ماند بلكه بيشتر به اشكال پيوسته در مي آيد اندول استيك اسيد و گلوتاميك اسيدوقندهايي مانند گلوكز و اينوزيتول براي اتصال به اكسين و تشكيل فرمهاي پيوسته به كار ميروند اتصال بين اسكوربيك اسيدواندول استيك اسيدنيز درگياه انجام مي شود انتقال اكسين در گياه در شكلهاي پيوسته صورت مي گيرد اكسين از محلي كه بيوسنتز شده است به ساير بخشهاي گياه منتقل و در آنجا ذخيره مي شود و يا آزاد شده و اثرات خود را آشكار مي سازد و يا اينكه پس از آزاد تجزيه و تخريب مي شود ميزان اكسين ها و فعاليت اين مواد در گياهان به بيوسنتز ، انتقال و متابوليسم آنها بستگي دارد اشكال پيوسته اكسين در مقايسه با اشكال آزاد اثرات فيزيولژيكي بسيار كمتري دارند .
جهت انتقال اكسين در گياه معمولا از طرف بالا به پايين است با بررسيهايي كه با استفاده از كربن راديواكتيو صورت گرفته است ديده شده است كه حركت اكسين ها عمدتا از بالا به طرف پايين مي باشد . حركات جانبي اكسين نيز مشاهده شده است و گزارشهاي كمي نيز مبني بر حركت از پايين به بالاي اكسين وجود دارد .
جهت انتقال اكسين در گياه درون سلولي است و از طريق آوندهاي آبكشي صورت مي گيرد ولي در ساير بافتها نيز در صورت زنده بودن سلولها اكسين جابجا مي شود. سرعت جابجايي اكسين در گياه بين 4 ميليمتر در ساعت تا 20 ميليمتر در ساعت گزارش شده است در غياب اكسيژن و حضور گاز كربنيك سرعت انتقال اكسين كاهش پيدا ميكند و در حضور اكسيژن سرعت اكسين افزايش مي يابداگر در اتمسفر گياه به جاي اكسيژن نيتروژن وجود داشته باشد سرعت جابجايي اكسين كاهش چنداني پيدا نكرده و متوقف نمي شود .
حركت هورمونهاي اكسين درگياه باعث شتاب در حركت هورمونهاي سيتوكينين و ژيبرلين مي گردد مهمترين ويژگي جابجايي اكسين انتقال قطبي آن است به اين معني كه اكسين از يك سو به سوي ديگر و از بالا به پايين حركت مي كند .
اگر قطعه اي از كولئوبتيل را به شيوه شكل 1 و به صورت الف و ب جدا كنيم به طوري كه الف قسمت بالايي و ب قسمت پايين آن باشد و قطعه كلئوپتينل را در همان جهتي كه در روي كولئوپتيل قرارداشته است ثابت كنيم قطعه آگار داراي اكسين را بر روي مقطع الف بگذاريم ودر قسمت ب قطعه آگار بدون اكسين را قرار دهيم پس از مدتي مشاهده مي كنيم كه آگار از قطعه بالايي به طقعه پاييني منتقل شده است .
حال اگر قطعه كولئوپتيل جدا شده را به طور عكس نسبت به وضعيت اصلي قراردهيم يعني مقطع الف در قسمت پايين و مقطع ب در بالا باشد قطعه آگار داراي اكسين را در بالا يعني بر روي مقطع ب و قطعه آگار بدون اكسين را در زير مقطع در پايين يعني در زير مقطع ب قرار دهيم مشاهده مي كنيم كه اكسين از قطعه آگار بالايي به آگار پاييني منتقل نمي شود
شكل1- نمايش حكت قطبي اكسين در گيا ه آگار داراي اكسين آگاربدون اكسين
با تجربيات متعددي كه بويژه با استفاده از مواد راديواكتيو صورت گرفته است نشخص شده است كه حركت اكسين يك طرفه بوده و حركت در ساقه ها از بالا به پايين صورت مي گيرد در حقيقت اكسين از محلي كه بيوسنتز مي شود به قسمتهاي ديگر گياه منتقل مي گردد جابجايي اكسين در ريشه ها به طور ضعيفي از پايين به بالا صورت
مي گيرد و سرانجام عواملي مانند اكسيژن ، گاز كربينك ، گرما ، فعاليتهاي متابوليكي و بازدارنده هاي متابوليكي در جابجايي اكسين موثرند. اين امر نشان مي دهد كه جابجايي قطبي اكسين در گياه وابسته به متابوليسم گياه است جابجايي اكسين در جهت جانبي و يا از پايين به بالا به صورت انتشار انجام مي گيرد.
در مورد انتقال قطبي اكسين نظريات مختلفي ارائه شده است.
ساختار و فيزيولژي ويژه آوندهاي آبكشي مي تواند عامل انتقال قطبي اكسين باشد بر اساس اين نظريه شاختمان نامتقارن را موثر در انتقال قطبي مي دانند وجود يك ميدان الكتريكي كه مربوط به تلمبه الكترون باشد ميز در انتقال قطبي موثر شناخته شده است بر اساس اين نظريه اكسين را يم آنيون در نظر مي گيرند كه مي تواند ميدان الكتريكي ايجاد كند اين نظريه چندان مورد قبول نيست زيرا با وارونه كردن كولئوپتيل پتانسيل معكوس مي شود در حالي كه چنين چيزي عملا مشاهده نمي گردد .
نظريه كيمواسمزي قطبيت بيشتر در مورد انتقال قطبي پذيرفته شده است اين نظريه در سال 1972 توسط روبري و شلدراك ارائه و در سال 1975 توسط راون تاييد گرديد بر اساس اين نظريه اكسين اسيدي ضعيف در نظر گرفته مي شود كه با لگاريتم آناليز 417 به IAA و H تجزيه مي شود با در نظر گرفتن اينكه سلولهاي آوندآبكشي در قطب پايين قابليت نفوذ بيشتري نسبت به IAA در مقايسه با قطب راسي دارند انتقال قطبي اكسين توسط اثر شيب PH و اختلاف پتانسيل اسمزي بين غشايي موجود مسيرهاي حركت اكسين را توجيه مي كند پيشنهاد دهندگان اين نظريه مهاجرت قطبي اكسين را تحت تاثير شيب PH و اختلاف پتانسيل بين غشائي بيان مي نمايند . متابوليسم گياه در اين جريان دخالت دارد به طوري كه بازدارنده هاي متابوليسمي مانند DWP و سانيدرين و اتيلن از حركات قطبي اكسين جلوگيري مي كنند (شكل 2)
شكل 2- اثر اتيلن با غلظت 25 ميكروليتر درليتر بر انتقال به سوي پايين اندول استيك اسيد نشان دار شده در رگبرگ اصلي برگهاي مركبات
تجزيه اكسين
اكسين در برابر نور خورشيد و اكسيژن تجزيه مي شود و اكر نور ضعيف باشد اكسين منتقل مي گردد در مراحل تجزيه اكسين تحت تاثير اكسيژن و نور نخست زنجيره كناري اكسين تجزيه مي شود و اندول آلدئيد توليد مي گردد .
گاهي اوقات تجزيه اكسين از زنجيره جانبي فراتر رفته و حلقه را نيز تجزيه مي كند .
اين تجزيه هاي اكسين تحت تاثير آنزيم پراكسيداز صورت مي گيرد كه يك هموپروتئين است براي اينكه تجزيه خوبي انجام شود امسيژن بايد به صورت O-O و اكسين نيز بايد قطبي باشد .
نخست اكسيژن وارد شده و فروپراكسداز را به فريك پراكسيداز تبديل مي كند و سپس اكسين معمولي به اكسين قطبي تبديل مي شود .
پس از آن فريك پراكسيداز بر روي اكسين قطبي مانند حالتي كه آنزيم بر سوبسترا اثر مي گذارد قرار مي گيرد .
شامل ورد 30 ص (تصاویر در سایت لود نشده اند اما در تحقیق اصلی موجودند)
مبلغ قابل پرداخت 36,000 تومان





