

اگر به یک وب سایت یا فروشگاه رایگان با فضای نامحدود و امکانات فراوان نیاز دارید بی درنگ دکمه زیر را کلیک نمایید.
ایجاد وب سایت یا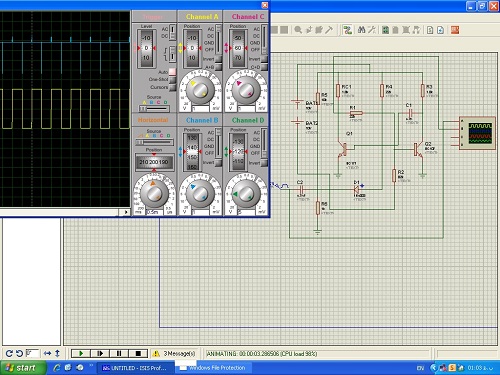 ازمایشگاه تکنیک پالس شبیهسازی با proteus
ازمایشگاه تکنیک پالس شبیهسازی با proteus تجزیه و تحلیل نقاشی مدرسه آتن
تجزیه و تحلیل نقاشی مدرسه آتن مقاله مدیریت سازمان های ورزشی
مقاله مدیریت سازمان های ورزشی كاربرد استخراج با سيال فوق بحراني در صنايع غذايي
كاربرد استخراج با سيال فوق بحراني در صنايع غذايي روش های نوین تجارت و تجارت الکترونیکی فرش دستباف
روش های نوین تجارت و تجارت الکترونیکی فرش دستباف تحقیق درباره ی Access
تحقیق درباره ی Access تحقیق كارشناسي ارشد عوامل موثر در روسپي گري فحشا
تحقیق كارشناسي ارشد عوامل موثر در روسپي گري فحشا هنر و گرافیک
هنر و گرافیک بررسی و طراحی نرم افزار حسابداری و مالی
بررسی و طراحی نرم افزار حسابداری و مالی تحقیق درباره ی هیدروژناسیون
تحقیق درباره ی هیدروژناسیون نقش و کاربرد انرژی هسته ای در کشاورزی
نقش و کاربرد انرژی هسته ای در کشاورزی تحقیق تنظيم كننده هاي رشد گياهي (هورمونهاي گياهي)
تحقیق تنظيم كننده هاي رشد گياهي (هورمونهاي گياهي) فروش فیلتر بورسی استریکلی فقط 75 هزار تومان
فروش فیلتر بورسی استریکلی فقط 75 هزار تومان کسب درآمد اینترنتی 300000 تومان در خانه در کمتر از 30 دقیقه
کسب درآمد اینترنتی 300000 تومان در خانه در کمتر از 30 دقیقه کسب درآمد روزانه حداقل یک میلیون تومان ! کاملا حلال و واقعـی !!
کسب درآمد روزانه حداقل یک میلیون تومان ! کاملا حلال و واقعـی !! مجموعه ی آموزش تعمیر لامپ کم مصرف (از مبتدی تا پیشرفته)
مجموعه ی آموزش تعمیر لامپ کم مصرف (از مبتدی تا پیشرفته) آموزش برنامه نویسی آردوینو
آموزش برنامه نویسی آردوینو کسب و کار اینترنتی در منزل
کسب و کار اینترنتی در منزل آموزش رایگان کسب درآمد از سایت الیمپ ترید ( olymp trade )
آموزش رایگان کسب درآمد از سایت الیمپ ترید ( olymp trade ) دانلود نمونه فاکتور آماده با فرمت ورد - اکسل و عکس
دانلود نمونه فاکتور آماده با فرمت ورد - اکسل و عکس آموزش ساخت بازی بدون دانش برنامه نویسی و طراحی سه بعدی مبتدی تا پیشرفته با نرم افزار
آموزش ساخت بازی بدون دانش برنامه نویسی و طراحی سه بعدی مبتدی تا پیشرفته با نرم افزار اموزش کسب درامد از اینترنت روزانه ۳میلیون تومان تضمینی و تست شده 1403
اموزش کسب درامد از اینترنت روزانه ۳میلیون تومان تضمینی و تست شده 1403 درامدزایی در خواب! (تعجب نکنید! بخوانید)
درامدزایی در خواب! (تعجب نکنید! بخوانید) مدار داخلی واکی تاکی(اموزش ساخت)
مدار داخلی واکی تاکی(اموزش ساخت) کتاب افزایش ممبر کانال تلگرام
کتاب افزایش ممبر کانال تلگرام دانلود100% رایگان نرم افزار تبلیغات در تلگرام + آموزش کامل و فیلم آموزشی
دانلود100% رایگان نرم افزار تبلیغات در تلگرام + آموزش کامل و فیلم آموزشی روش اصلی موفقیت در کنکور و آزمون ها(پزشکی، حقوق، مهندسی، نمونه و تیزهوشان) با پکیج کنکورپلاس
روش اصلی موفقیت در کنکور و آزمون ها(پزشکی، حقوق، مهندسی، نمونه و تیزهوشان) با پکیج کنکورپلاس
مولكول و اكسيژن ريشه واكنشهاي آن: 4
منابع بيولوژيكي اكسيژن واكنشدار: 6
تشكيل اكسيژن واكنشدار در كلروپلاست گياهان: 7
توليد اكسيژن واكنش دار در ديگر بخشهاي سلولي: 10
دفاع بر عليه اكسيداتيو حاصل از استرس 11
آنزيم سوپر اكسيد دسيموتاز (SOD) : 12
اسكوربات پراكسيداز، گلوتاتيون ردوكتاز و مونوديهيدوكسي اسكوربات ردوكتاز: 13
كاتالاز 15
پروتئين و آنزيمهاي اضافي: 16
آنتي اكسيدانهاي غير آنزيمي: 17
ويتامينE (tocopherol): 17
كاروتنوئيدها: 18
گلوتاتيون: 18
تركيبهاي گوناگون: 19
PHOTOINHIBITION 19
تركيبات TRIAZOLE ، سم ها و HERBICIDES : 20
مواد سمي: 21
آلودگي هوا: 22
استرس نمك: 23
استرس خشكي و گرما: 24
سرما و يخ زدن: 24
مقدمه: 25
استرس اكسيداتيو و فتوسنتز: 28
تنظيم زنجيزه انتقال الكترون فتوسنتز: 29
Photoinhibition : 33
انتقال الكترون به اكسيژن: واكنش mehler : 35
آنزيمهاي آنتي اكسيداني در كلروپلاست: 38
نقش گلوتاين در سيكل مجرد اسكوربات در كلرو پلاست: 40
پاسخهاي استرس محيطي و محدوديت ها: 41
رابطه بين كلو پلاست و ديگر قسمتهاي سلول: 43
تنفس نوري: 44
بخش بندي اسكوربات: 45
نقش پراكسيد و ساليسيك اسيد: 48
نقش سوپرسوپر اكسيد درون سلولي در hr: 48
تشكيل تومور در گياهان: 48
نيتريك اكسيد و استرسي گياهان: 49
برهم كنش آفت گياهان و آنفجار اكسيداتيو: 49
تنظيمي از پاسخ ها را استرس اكسيداتيو: 50
تنظيم سيستم دفاعي د ر گياهان: 50
اتم اكسيژن فراوان ترين عنصر در پوسته زمين است و در اتمسفر و آب براي شكلهاي هوازي حيات مورد نياز مي باشد مخزن اكسيژن كره زمين نتيجه ساخته شدن آن از واكنشهايي مانند فتو سنتز است 37 EMOL و 1 EMOL=108 MOLES. فتوسنتز واكنشي است كه در آن دي اكسيژن (2O) از آب آزاد ميشود دي اكسيژن تقريبا بطور دائمي در تنفس استفاده ميشود و بيشتر از آن جهت استفاده ميشود كه پذيرنده نهايي الكترون است. اتم اكسيژن در آنواع مولكولهاي آلي بوسيله تنوعي از واكنشهاي آنزيمي (مانند اكسيژن ساز) و غير آنزيمي ثابت ميشود.
موجودات بي هوازي هر چند از عهده اثر مخالف اكسيژن بر مي آيند. در غلظت بالاتر از اكسيژن اتمسفر، دي اكسيژن ممكن است (2O) بازدارنده يا غير فعال كننده آنزيم معيني باشد و يا اينكه ممكن است با CO2براي ثابت شدن بوسيله 1، 5 – بيسفسفات كربوكسيلاز اكسيژناز رقابت كند كه اين باعث ميشود كه ارزش انرژيتيك فتوسنتز افزايش يابد. بطور كلي اثر سمي اكسيژن به طور عمده توسط مشتقات واكنشي (واكنش پذير) آن اعمال ميشود، در حاليكه دي اكسيژن در حالت پايدار اتم نسبتا غير قابل واكنش است و مي تواند در حالت آرامش همراه مواد الي وجود داشته باشد اجباراتي جزئي از ساختار آنها گردد. اين ويژگي توسط چرخشهاي موازي دو الكترون جفت نشده دي اكسيژن، كه داراي يك سدانرژي براي واكنش اكسيژن با تركيبات غير راديكال است (ممانعت چرخش) نشان داده ميشود.
براي اينكه اكسيژن به طور شيميايي واكنش پذير باشد بايد بطور فيزيكي يا شيميايي فعال شود. فعاليت فيزيكي بطور عمده توسط انتقال انرژي تحريك از يك رنگيزه فعال شده با نور همانند كلروفيل تحريك شده به اكسيژن اتفاق ميافتد با جذب انرژي كافي چرخش يك الكترون معكوس ميشود. اولين حالت منفرد از اكسيژن (به صورت O2يا 2 نشان داده ميشود) يك نوع واكنش پذير متداول است. اين حالت اكسيژن قابليت انتشار زيادي دارد و قادر به واكنش با ملكولهاي الي است (كه الكترونها معمولا جفت شده هستند) و به غشاهاي فتوسنتزي آسيب ميرسانند.
فعاليت شيميايي مكانيزم ديگري براي ممانعت چرخش الكترون و فعال كردن است. اين عمل توسط احيا يوني والنت (يك ظرفيتي)دي اكسيژن با افزايش الكترونهاي يكي توسط ديگري آنجام مي گيرد. چهار الكترون و چهار پروتون براي احيا كامل اكسيژن به آب نياز است. همه سه حد واسط احيا يك ظرفيتي(univatent) مثلا سوپر اكسيد O2o. پراكسيد هيدروژن (H2o2) راديكال هيدروكسيل OHo از نظر شيميايي فعالند و از نظر فيزيولوژيكي سمي هستند اين سميت به وسيله نيمه عمرها ي كوتاه آنها قبل از واكنش با تركيبات سلولي و در مقايسه با نيمه عمر دياكسيژن انعكاس داده ميشود، (بيشتر از 100 ثانيه جدول 1) نوع اكسيژن واكنشي كه با يك ملكول الي برخورد ميكند يك الكترون را از آن خارج ميكند و در يك واكنش زنجيره اي به صورت راديكالهاي پراكسيل(Rooo)و آلكوكسيل (Roo) درآيد.
سوپر اكسيد اولين توليد احيا شده از حالت بنيادي اكسيژن است كه اين توانايي را دارد هم از اكسيد شدن و هم از احيا شدن ايجاد شود اين ماده با چند ماده توليدي از واكنشهاي ديگر ممكن است واكنش دهد كه اين عمل بطور خود به خود و يا بوسيله آنزيمهاي جهش نيافته منجر به توليد H2o2 ميشود.
پراكسيد هيدروژن يك راديكال آزاد نبوده اما بعنوان اكسيد كننده و عامل احيا كننده در تعدادي از واكنشهاي درون سلولي شركت ميكند بر خلاف سوپر اكسيد، H2o2 انتشار بيشتري از ميان غشاها و بخشهاي كردهبندي شده سلول داشته و ممكن است مستقيما آنزيمهاي حساس را در غلظت كم غير فعال كند مانند سوپر اكسيد، H2o2 پايداري بيشتري دارد. بنابراين از ديگر آنواع آن اكسيژنهاي واكنش اثر سمي كمتري دارد تهديد عمده سوپر اكسيد و H2o2 در توانايي آنها به توليد زياد راديكالهاي هيدروكسيل واكنشدار ميباشد.
راديكالهاي هيدروكسيل از آنواع اكسيد كننده قوي در سيستم بيولوژيكي بشمار ميروند. اين راديكال ها بطور غير ويژه اي با هيچ مولكول بيولوژيكي واكنش نمي دهند اين موضوع به انتشار آن بر ميگردد كه انتشار آن در درون سلول به آندازه قطر 2 مولكول از جايگاه توليد آنها انجام ميگيرد.
هيچ نوع تميز كننده اي كه بتواند OHoرا جمع آوري كند شناخته نشده است. اگر چه پيشنهاد شده است كه چندين متابوليت همانند اوره يا گلوكز در سيستمهاي جانوري جمع كننده OHo هستند جديدا هم نقشي براي OHo در متابوليسم پلي ساكاريدهاي ديواره سلولي پيشنهاد شده است نوع واكنشهاي مختلفي كه در بالا شرح داده شده است به سبب تغييرات: 1- جلوگيري از آنزيمهاي حساس 2- كاهش كلروفيل يا بي رنگ شدن 3- پراكيداسيون ليپيدها مي باشد: به دليل يورش راديكالهاي آزاد، H2o2 و اكسيژن منفرد به اسيدهاي چرب غير اشباع ليپيدهاي هيدروپراكسيد توليد ميشود و در حضور مواد كاتاليزي راديكالهاي الكوكسيل (alkoxyl) و پراكسيل (peroxyl) به زنجيره واكنش در غشا سلولي انتقال مي يابند و ليپيدهاي ساختاري و غشاهاي سازمان يافته و بي عيب تغيير و تجزيه مي شوند، بعلاوه تعدادي الدئيد و هيدروكربنهاي توليد شده بوسيله پراكيداسيداسيون باعث اثر سمي در سيستمهاي جانوري ميشود
4- حمله ناشي از عدم تشخيص بوسيله راديكال ها به ملكولهاي الي مانند DNA . تغيرات زيادي در اثر حمله OHo به DNA ايجاد ميشود كه شامل شكست رشته ها كه ممكن است دوباره جفت شدن آنها را دچار مشكل كند و يا واكنشهاي متناوب پايه اي باشد. پروتئينهايي كه د ر معرض OHo قرار مي گيرند تغييرات بر جسته اي پيدا مي كنند كه شامل تغيير توالي آمينو اسيدهاي ويژه، كه شامل تغيير متوالي آمينو اسيدهاي ويژه، قطعه قطعه شدن پلي پپتيد، اجتماع و دناتوره شدن پروتئينها و آمادگي آنها براي تجزيه مي باشد.
در همه جريانهاي متابوليكي طبيعي در موجودات هوازي آنواع اكسيژن واكنشدار ( ROS) تشكيل ميشود در شكل 1 تعدادي از مسيرهاي فيزيولوژيكي گياه كه توليد اكسيژن واكنش دار مي كنند مورد بحث قرار مي گيرند اگر چه تعدادي از راديكالهاي اكسيژن كه در اثر اشريشيا هوازي ساخته مي شوند زودتر كشف شدهاند در رشد هوازي E.coli راديكال O2oدر طول انتقال الكترون در غشا توليد ميشود فقط 4% درصد از الكترونها انتشار يافتند و مكانيسمي نياز است تا از تجمع سوپر اكسيد جلوگيري كند از نمونه موتانهاي E.coli غشاهاي تهيه شده و سطح سوپراكسيد آندازه گيري شده كه تهي از سوپراكسيد دسيموتازبود sodA) (sodBچنين موتانهايي رشد كمي را نشان دادند و از طريق چندين موجود اگزوتروف ثابت گرديد كه soD (سوپر اكسيد اسيموتاز) نقش حياتي در كاهش سطح پايدار o2o به ميزان دارد يكي از بيشترين آنزيمهايي كه در سيستم E.coli به سوپر اكسيد حساس مي باشد آنزيم اكونيتازميباشد كه در چرخه TCA نقش دارد غير فعال شدن اكونيتاز ميباشد كه در برگشت پذيري اكيداتيو بوسيله O2o آنجام مي گيرد و پيشنهاد شده است كه با ايجاد يك نقش تدافعي [ ciruit breaking ] باعث توليد NADPH در راه كوتاهتر ميشود.
كلرو پلاستها منابع بزرگي از تشكيل اكسيژن واكنش دار در گياهان هستند. در انرژي نوري زياد كارايي آنها بالا رفته و ايجاد مواد احياكننده همانند NADPH ميكند. چندين مسير يا جايگاه وابسته فعاليت اكسيژن در كلرو پلاست وجود دارد كه در مواقعي به سمت توليد اكسيژنهاي واكنشي هدايت ميشوند. اهميت زياد از اين جهت است كه سمتي از PSI احياكننده است كه يك الكترون ممكن است بوسيله كارير (حامل) غشايي به o2 انتقال يابد (واكنشmehler )، در عوض از جريان الكترون به سمت پائين در نهايت NADP+احيا ميشود (شكل 1) تحت وضعيتهايي كه قابليت استفاده پذيرندههاي الكترون از PSI محدود مي شوند به عنوان مثال وقتي كه سيكل كالوين به آندازه كافي و با سرعت NADPH مصرف نمي كند سوپر اكسيد در غشا شكل ميگيرد.
درPH پائين سوپر اكسيد خود به خود از انتشار زياد H2O2 جلوگيري ميكند، به طور ديگر سوپر اكسيد ممكن است با پلاستو سيانين يا سيتوكروم F واكنش داده و آنها را احيا كند، نتيجه واسطه اي بودن سوپراكسيد باعث ميشود كه سيكل الكترون در اطراف PSI جريان يابد اين مكانيسم پيشنهاد ميكند كه توليد سوپر اكسيد يك نقش تنظيم كنندگي دارد كه با تغيير دادن يا چرخش بيش از حد الكترون د رهمان زمان مانع انتشار راديكالها به يك صرف از غشا ميشود.
پراكسيد هيدروژن از بيشترين توليدات كلروپلاست است كه به طور بي تناسبي از سوپر اكسيد و بوسيله آنزيم SOD ايجاد ميگردد، كه دراثر از بين رفتن خود بخود بيثباتتر ميشود. از منابع ديگر توليد H2O2 تنفس نوري است كه بوسيله فعاليت اكسيژنازي آنزيم 1، 5 ريبو لوز بيس فسفات كربوكسيلا زد راستروماي كلروپلاست شروع ميشود و در نتيجه د رپراكسي زوم توليد H2O2ميشود (شكل 1).
تنفس نوري ممكن است يك مكانيسم محافظتي براي بازسازي پذيرنده الكترون باشد كه با اين عمل اجازه مي دهد كه جريان ميدهد كه جريان الكترونهاي فتوسنتزي تحت وضعيتهاي تثبيت كم كربني ادامه يابد.
در مقايسه با چرخه الكترون اطراف PSI، سيكل تنفس نوري باعث از هم پاشيدگي ATP و NADPH ميشوند. يك مكانيسم مهم از تنظيم كه باعث جفت شدن تثبيت كربن در استروما به جريان الكترون فتوسنتزي ميشود. اين است كه وقتي انتقال دهندههاي الكترون در واكنشهاي نوري اكسيد مي شوند آنزيمهاي سيكل كالوين بطور برگشت پذيري غير فعال وقت انتقال دهندهها احيا شدند. اين آنزيم ها دوباره فعال مي شوند چنين تنظيمي بوسيله واسطههايي مانند تيورود كسين،PH استروما و ديگر فاكتورها آنجام ميگيرد.
پراكسيد هيدروژن از طريق اكسيد كردن گروههاي تيول باعث مختل شدن كار اين مكانيسمهاي حفاظتي مي شوند همچنين پراكسيد هيدروژن باعث غير فعال شدن آنزيمهاي چرخه كالوين بطور غير بازگشتپذير ميشود.
بنابراين غلظت آن بايد در كلروپلاست پايين نگه داشته شود همچنين آن ممكن است آنزيم SOD مس / روي غير فعال كند. راديكالهاي هيدروكسيل ممكن است در طي واكنشهاي كاتاليزي در تمام سلولهاي زنده بوسيله تغيير يونهاي آهن و مس وقتي كه H2O2 و سوپر اكسيد حضور دارند تشكيل شود:
1- واكنش Fenton كه در نتيجه آن، OHo از H2O2 توليد ميشود.
2-يون آهن غيرچرخهاي همانند يك احيا كننده فعال بوده و با اضافه شدن سوپر اكسيد به واكنش 1 بصورت زير ادامه مييابد.
3- حاصل جمع واكنش 1 و 2 كه واكنش haber-weiss ناميده ميشود.
SOD, O2o با هم يك منبع تميز كننده مشترك ا زH2o2 از بين مي رود، علاوه بر آن آنواع اكسيژن واكنشدار ديگري را نيز از بين مي برند خطر اصلي پراكسيد هيدروژن و سوپراكسيد غير مستقيم بوده و آن وقتي اتفاق مي افتد كه آنها اجازه پيدا ميكنند كه د رهمان جايگاههاي سلولي تجمع يابند و متابوليسم آهن و مس د ر تماس با استرس اكسيداتيو هستند مطابق يك گزارش چالش آنگيز بوسيله yim و همكارانش (1990)، راديكالهاي هيدروكسيل بوسيله آنزيمcu/zn SOD بصورت محلول آزاد ميشود كه خود با آن واكنش داده و توليد پراكسيد هيدروژن ميكند. اكسيژن منفرد وقتي در كلروپلاست شكل ميگيرد كلروفيل برانگيخته از نور در حالت سه گانه با دياكسيژن واكنش مي دهد. دوباره وقتي كه ATP و NADPH بوسيله واكنشهاي چرخه كالوين كمتر استفاده مي شوند ميزان آن زيادتر ميشود تعدادي از فاكتورهاي استرس كه محدود كننده همانند سازي CO2هستند ممكن است باعث افزايش اين جريان ها شود (مانند بسته شدن روزنه ها). انرژي باقي مانده بر انگيخته از اينرو از طريق فلورسانس يا فروكش كردن كارتنوئيدها پراكنده ميشود. بعلاوه اكسيژن منفرد ممكن است بوسيله پراكسيدازهاي معين در گياهان توليد شود.
1- ميتو كندري:
ميتوكندري اكسيژن را در طول انتقال الكترون تنفسي مصرف ميكند جايگاههاي مختلفي از نشت الكترون و رهاسازي سوپراكسيد و پراكسيد هيدروژن د ر تنفس پيشنهاد شده است يكي از اين جايگاههاي ويژه جايگاه اكسيد از متناوب حساس به سيانيد است كه در ميتو كندري وجود دارد.
2- شبكه آندوپلاسمي، پراكسيزوم و گلي اكسيزم:
چندين واكنش اكسيداتيو مختلف از شبكه آندو پلاسمي صاف و ميكروزوم ها سر چشمه مي گيرد.
اكسيژنازهاي داراي كارهاي مختلف و مخلوط مانند سيتوكروم P450 واكنش مهم هيدروكسيلاسيون را در مسير موالونيك اسيد كه اضافه كردن اتمهاي اكسيژن به مولكولهاي سوبسترا است را انجام ميدهند.
NAD (P) H دهنده الكترون است و ممكن است بوسيله چنين واكنشهايي سوپر اكسيد رها شود. پراكسي زوم و گلي اكسيزوم اندامكهاي تك غشايي هستند كه آنزيمهايي كه در بتا اكسيداسيون اسد چرت و تركيبات دو كربنه سيكل تنفس نوري شركت دارند را شامل مي شوند كه در آن آنزيمهاي گلي كولات اكسيدار الكترون را از گلي كولات به اكسيژن انتقال داده و توليد H2O2 ميكند. گزانتين اكسيداز، اورات اكسيداز، NADH اكسيداز از سوپر اكسيدازهاي عمومي هستند
3- غشا سلولي و قسمت بندي اپوپلاست:
NAD(P) H اكسيد از جزئي است كه در همه جان غشا پلاسمايي وجود دارد و ممكن است توليدH2O2 و سوپر اكسيد كنند. اكسيژن واكنشي ممكن است در اپوپلاست ايجاد شوند. آپوپلاست اولين جايگاه حمله پاتوژن است، آن شامل يكي از اولين خطهاي واكنش دفاعي گياهان است و با اين عمل آپوپلاست نيز در گير اكسيژن واكنشي است. بيشتر مسير بيوسنتز مشترك در آيوپلاست بيوسنتز ليگنين است جايي كه Phenypropanoid بعنوان پيش رو از ليگنين در شروع واكنش بوسيله پراكسيداز در جهت ديگر توليد H2O2 ميكند.
در ديواره سلولي NADH نياز است كه بوسيله مالات دهيدروژناز ديواره سلولي تامين ميشود و شايد بوسيله يك NADH اكسيداز به شكل H2O2مورد استفاده قرار ميگيرد.
آمينواكسيدازها اكسيژن فعال شده را بوسيله يك NADH اكسيداز به شكل H2O2مورد استفاده قرار مي گيرد. آمينو اكسيدازها اكسيژن فعال شده را بوسيله احيا ديآمينها يا پليآمينها به كينون تبديل مي كنند كه همراه با تشكيل پراكسيد ميباشد.
قبلا در باره اجزا ويژه محافظتي در گياهان از اثر اكسيژن واكنشي بحث شد. ما بايد توجه داشته باشيم كه مكانيسمهايي عالي انتقال الكترون را تنظيم ميكند و فتو سنتز در كل اولين دفاع بر عليه استرس اكسيداتيو است. اين مكانيسم ها مسئولهاي جفت شدن خوب واكنشهاي نوري و تاريك فتوسنتز هستند و باعث سازگاري سيستم نوري فتوسنتز در برابر تغيير وضعيت ميشود. از تحريك پذيري زياد و توليد زياد سيستم فتوسنتزي اجتناب ميشود و اكسيژن واكنشي در حد مينيمم احيا ميشود.
با وجود اين ما شناختيم كه به تدريج و پيوسته آنواع اكسيژن فعال در همه قسمتبنديهاي سلولي و به عنوان يك محصول فرعي از متابوليسم ها نرمال سلولي توليد ميشود كه نشان مي دهد بقا سلولي وابسته به حمايت كافي است. همه شكلهاي هوازي زندگي شامل چندين خط دفاعي هستند كه شامل آنزيمهاي جمع كننده (پاك كننده) و آنتي اكسيدانهاي غير آنزيمي هستند چنين گوناگوني دفاعي نيازمند مي باشد چون:
1- انواع اكسيژن واكنش (ROS) در قسمت بندي سلول و خارج سلولي توليد ميشوند .
انواع واكنش دار خاصيت مختلفي همانند توانايي انتشار، توانايي حل شدن و ميل تركيبي با مولكولهاي بيولوژيكي مختلف را دارند بنابراين يك دستگاه مختلف از دفاع ملكولي در فاز غشايي و غاز مايع نياز است كه در آنجا فعاليت كند تا فورا راديكالهايي كه در آنها شكل گرفته آند را غير فعال كنند.
شامل ورد 50صفحه ای میباشد
مبلغ قابل پرداخت 22,000 تومان





